Fagfellevurdert artikkel viser at embryonal variasjon hos virveldyr motsier felles avstamning
Av Casey Luskin; 23. mars 2023. Oversatt herfra
Evolusjonsbiologer hevder ofte at embryoer fra virveldyr utvikler seg på svært like måter, noe som gjenspeiler deres felles aner. Men en fagfellevurdert artikkel i BIO-Complexity, av David Swift (forfatter av Evolution Under the Microscope), gir en innsiktsfull gjennomgang om emnet. Artikkelen har tittelen "Den mangfoldige tidlige embryonale utviklingen av virveldyr og implikasjoner angående deres aner." -lenke; Swift viser at, til tross for vanlige påstander, utvikler ikke virveldyr seg likt, i henhold til spådommene om evolusjon.
 Bilde 1. Kyllingembryo
Bilde 1. Kyllingembryo
Han åpner med å fremme sin tese:
Det er velkjent at den embryonale utviklingen av virveldyr fra forskjellige klasser (f.eks. fisk, reptiler, pattedyr) går gjennom et "fylotypisk stadium" når de ser like ut, og denne tilsynelatende homologien blir mye sett på som bevis på deres felles aner. Til tross for deres morfologiske likheter, og i motsetning til evolusjonære forventninger, oppstår de fylotypiske stadiene til forskjellige virveldyrklasser på radikalt forskjellige måter. Dette mangfoldet motvirker klart det overfladiske utseendet til homologi i det fylotypiske stadiet, og den enkle slutningen er at virveldyr ikke har utviklet seg fra en felles vertebrat-forfedre. Mangfoldet strekker seg gjennom alle stadier av tidlig utvikling - inkludert spaltning og dannelse av blastula, gastrulering, nervedannelse og dannelse av tarm og ekstra-embryonale membraner.
Nå krever ikke intelligent design at felles avstamning er feilaktig, men selv en styrt form for felles avstamning, kan føre til forskjellige forutsigelser fra strengt ikke-styrt avstamning med modifikasjon. Dermed fant Darwin og påfølgende evolusjonister som Ernst Haeckel at embryologi var en avgjørende bevislinje som støtter Darwins avhandling. I følge Swift forutsier felles aner at virveldyr-utvikling bør vise slående homologier. Men mer enn det, "Hvis felles aner er forklaringen på homologier, bør ikke bare homologe organer være avledet fra tilsvarende embryonale vev (kardinalkriteriet for homologi), men de bør også utvikles ved sammenlignbare prosesser." {understreking og kursiv ved oversetter}
Svært forskjellige veier
Kjernen i Swifts avhandling er at selv om virveldyr-embryoer passerer gjennom et lignende "fylotypisk stadium", er utviklingsveiene svært forskjellige:
Til tross for deres morfologiske likheter, og i motsetning til evolusjonære forventninger, er det et slående faktum at de "fylotypiske stadiene" til forskjellige grupper av virveldyr oppstår på bemerkelsesverdig ulike måter, selv med nøkkelvev som kimlagene (se nedenfor), som stammer fra fullstendig forskjellige tidlige embryonale kilder. Disse observasjonene tilbakeviser klart den antatte evolusjonære homologien til det fylotypiske stadiet av virveldyr, og undergraver derfor slutningen om felles aner basert på den antatte homologien. [Utheving i originalen.]
Han trykker opp igjen "timeglassmodellen" for utvikling av virveldyr og påpeker at dette bare er en 'observasjon' om utvikling - ikke en forklaring på hvordan den oppsto: Et av nøkkelstadiene i utviklingen som fører til dette lignende 'fylotypiske stadiet' er gastrulasjon , som Swift bemerker er avgjørende, fordi det etablerer den grunnleggende kroppsplanen og "fører til etableringen av kimlagene - ektoderm, mesoderm og endoderm - som alt kroppens vev er avledet fra."
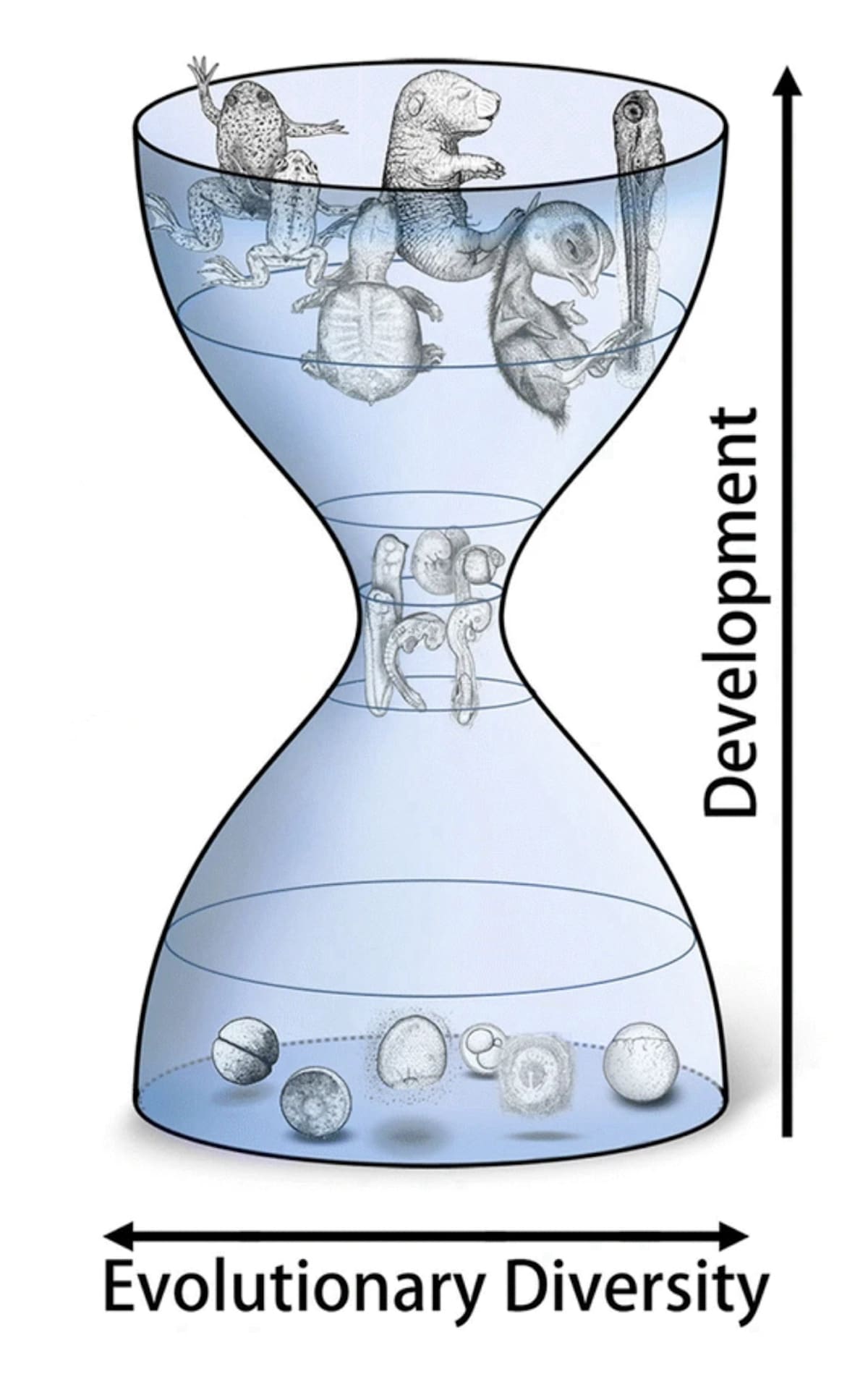 Bilde 2. Timeglasset
Bilde 2. Timeglasset
Forskjeller i utvikling av virveldyr
Swift siterer forskjellige spesifikke forskjeller i virveldyrutvikling for å utvikle sin sak, spesielt i gastrulering. Han forutsier at "fra et evolusjonært perspektiv ville vi helt sikkert forvente at gastrulering er 'bevart'", men finner at "for nesten alle hovedklassene av virveldyr" er det viktige forskjeller i gastrulering, inkludert "gasstrulasjonsmekanismen, er betydelig forskjellig fra noen av de andre," og "kildevevet til kimlagene er forskjellige."
Etter å ha gjennomgått mekanismer for gastrulasjon i forskjellige klasser av virveldyr, bemerker Swift at "det store utvalget av strukturer i blastulaene til forskjellige klasser av vertebrater utfordrer synet om at det resulterende embryonale vevet kan betraktes som ekvivalente eller homologe." Nærmere bestemt, hos forskjellige typer virveldyr blir forskjellige deler av blastulaen til slutt selve embryoet. Han beskriver disse forskjellene som følger:
*Chondrichthyans (ryggstrengdyr): "Det er et encellet tykt epitellag som danner den øvre overflaten av blastula."
*Teleoster (benfisk, for eksempel sebrafisk): "Det er et flercellet lag, under det overliggende omsluttende laget."
*Amfibier: "Det er hele blastula, som omfatter den flerlagede kuppelen på den øvre halvkulen og cellemassen i den nedre halvkule."
*Reptiler og fugler: "Det er den øvre overflaten av blastulaen, som består av et encellet tykt epitellag som ligger over hypoblasten."
*Placentale pattedyr (f.eks. primater): "Det er del av den indre cellemassen, innenfor den ytre trofoblasten."
Han siterer ytterligere forskjeller mellom hvilke celler som blir endodermen og hvilke som blir mesodermen, og bemerker:
*i amnioter (krypdyr, fugler, pattedyr) oppstår celler som er internalisert fra et sentralt område av epiblasten, dvs. den presumptive endoderm og mesoderm er omgitt av presumptiv ektoderm; mens
*hos anamnioter (kondrichthyaner, teleoster, amfibier) er cellene som internaliserer fra kanten av epiblasten, dvs. den presumptive endodermen og mesodermen omgir den presumptive ektodermen.
Swift oppsummerer store forskjeller i gastrulasjonsmekanismene som følger:
*Chondrichthyans: ved at celler ruller over et bakre overheng av epiblasten.
*Teleoster: ved involusjon rundt kantene av epiblasten når den sprer seg rundt eggeplommen.
*Amfibier: ved involusjon gjennom en ringformet blastopore.
*Reptiler: ved involusjon gjennom en kanallignende blastopore.
*Fugler: ved at celler trenger inn gjennom en primitiv stripe, dannelsen av den primitive stripen blir ledsaget av vekst av en underliggende endoblast.
*Placentale pattedyr: ved at celler trenger inn gjennom en primitiv stripe.
"Tre vesentlige forskjeller"
Swift finner dermed at disse forskjellige utviklingsmåtene ikke kan betraktes som homologe:
I lys av disse tre vesentlige distinksjonene - den forskjellige overordnede strukturen til blastulaene, de forskjellige delene av blastulaen som blir embryoet, og de forskjellige relative posisjonene til den presumptive ektodermen og mesoderm/endoderm i fostervann og anamnioter - er det ingen tvil at vevene som blir embryoet ikke er ekvivalente, og dermed er langt fra å være homologe på tvers av de 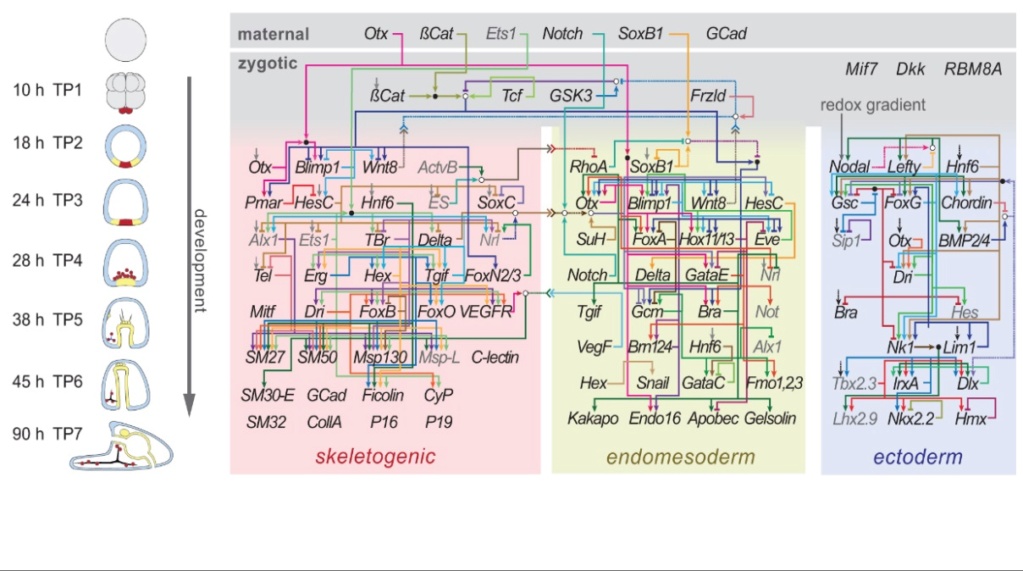 ulike virveldyrklassene. {oversetters understreking}
ulike virveldyrklassene. {oversetters understreking}
Bilde 3. Skjematisk utvikling av ulike kimlag
I følge Swift antyder disse grunnleggende forskjellene i tidlige vertebrat-utviklingsmekanismer under gastrulering, at virveldyr ikke deler felles avstamning:
-Den enkle konklusjonen Å trekke fra dette radikale mangfoldet av deres tidlige embryonale utvikling, er at det viser at virveldyrene ikke har utviklet seg fra felles virveldyr-forfedre. Denne konklusjonen kan bare unngås, hvis det er troverdige forklaringer på hvordan et slikt mangfold av tidlig utvikling kan ha oppstått fra utviklingen som råder i en felles stamfar (enten de er like dagens cefalochordater, eller ikke) på en evolusjonær måte, via endringer som (i) hadde en realistisk sannsynlighet for å inntreffe, (ii) opprettholdt levedyktighet og (iii) i de fleste tilfeller, tilbød en betydelig fordel, som kunne favoriseres av naturlig utvalg.
Å møte disse evolusjonære kravene utgjør imidlertid en stor utfordring. Swift siterer en fremtredende utviklingsbiolog, Rudolf Raff, som skrev: "Man kan med rimelighet forvente at mekanismer for tidlig utvikling er spesielt motstandsdyktige mot modifikasjoner, fordi all påfølgende utvikling kommer fra tidlige prosesser." Swift kaller dette en "sunn konklusjon" fordi kompleksiteten i utviklingen av virveldyr krever at mange koordinerte modifikasjoner vil være nødvendige for å fundamentalt endre hvordan utviklingen fortsatte. Han finner dermed at "på grunn av den gjensidige avhengigheten av mekanismene som er involvert, må konstruktive endringer i embryonal utvikling innebære koordinert produksjon av og/eller endringer i flere gener, for eksempel for transkripsjonsfaktorer og DNA-sekvensene de virker på, som er uoverkommelig usannsynlig."
Dette fører til et "ventetidsproblem" der flere koordinerte endringer vil være nødvendig for å gå over fra ett utviklingsregime til et annet, og utgjøre "en formidabel utfordring for antatte evolusjonære scenarier," ettersom å generere disse endringene ville være "generelt langt over den tilgjengelige tiden." Se evt. ventetidsproblematikk her og -her.
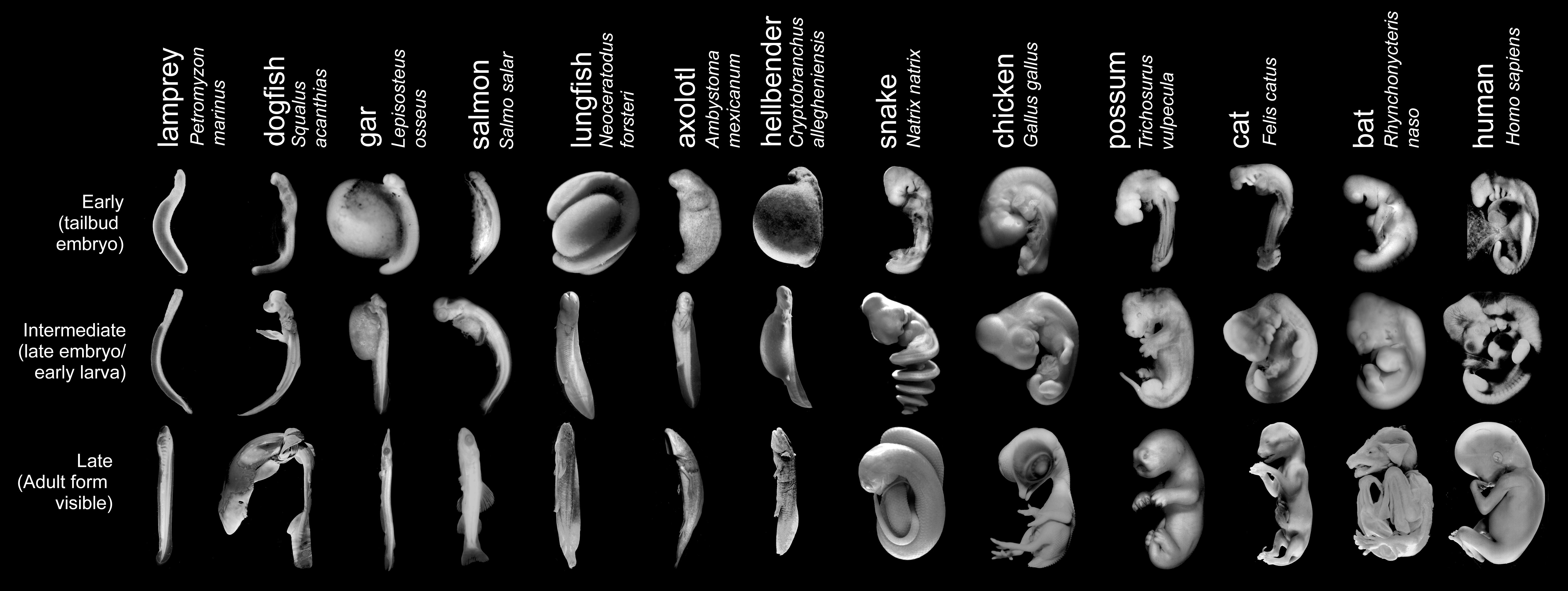
Bilde 4. Foto av embryo-utvikling (3 ulike stadier)
Casey Luskin
Assisterende direktør, senter for vitenskap og kultur
Casey Luskin
Luskin er geolog og advokat med høyere grad i vitenskap og jus, som gir ham ekspertise både i de vitenskapelige og juridiske dimensjonene av debatten om evolusjon. Han fikk sin doktorgrad i geologi fra University of Johannesburg, og BS- og MS-grader i geovitenskap fra University of California, San Diego, hvor han studerte evolusjon på både hoved- og lavere nivå. Hans jusgrad er fra University of San Diego, hvor han fokuserte studiene på første Amendment, utdanningslov og miljørett.
Oversettelse ved Google-oversetter, sett gjennom av Asbjørn E. Lund